



By Dr. Thomas T. Yamashita
Why are some microorganisms harmless and others pathogenic (disease-causing)?
The basic characteristics of the ancestral microorganism of millions of years gone past is for the most part retained in the majority of present day forms. It is a principle of developmental biology that the shift towards variations of forms—greater diversity—is governed by the unique environment and the variable conditions surrounding the microorganism in its procurement of food. In the struggle for survival and with ever-increasing competition were found the selective pressures to weed out forms less adapted for the various, intricate requirements of that competitive environment. Pathogenic forms of microorganisms—referred to as pathogens—are generally specialized, with a unique set of enzymes which allow them to enter the host tissues, thereby minimizing competition and allowing it to secure sustenance unavailable to more general, non-infectious organisms.
Are some pathogenic organisms more pathogenic than others?
Pathogenic microorganisms can be divided into two major groups:
- Facultative pathogens
- Obligate pathogens
The facultative pathogen normally exists as an organism which lives on debris or on aging/dying tissues, much as mushrooms and other fungi do. Under certain conditions, the facultative pathogen (which literally translates to “conditional pathogen”) can gain a foothold in the host and become pathogenic (such as in hosts with impaired immune responses).
The obligate pathogen has developed a long-standing relationship with its host over eons and eons of time. It represents the ultimate developmental specialization in a pathogen, as the obligate pathogen has lost its ability to live outside the host. As opposed to the facultative form, the obligate pathogen’s enzyme system is geared almost entirely to the host, most being near impossible to culture on laboratory media.
In short, facultative pathogens can cause infection when the opportunity arises, allowing them to survive and reproduce more easily, while obligate pathogens must cause infection in order to survive and reproduce.
What is the most common form of plant pathogen?
Over 95% of all plant pathogens fall into the classification of “facultative pathogens.” Their presence in almost every environment is facilitated by their ability to adapt to a broad spectrum of substrates. Further, they are able to survive outside the host, sustaining on debris or aging or dying host tissues. Many of these facultative pathogens have also developed survival mechanisms or “overwintering biology” endowing them with an ability to remain viable in the absence of a host. A few of these common facultative forms are: Botrytis spp., Cladosporium spp., Monilinia spp., Sclerotinia spp., Cytospora spp., Ceratocystis spp., Pythium spp., Phytophthora spp., Fusarium spp., Verticillium spp., Botryosphaeria spp., Armillaria spp., Pseudomonas spp., Erwinia spp. and many, many others.
Why is it that soil-borne diseases tend to “creep up” on farmers over time?
Most soil-borne pathogenic organisms are not well adapted to the microbially rich, active soils in which crops are grown. When the populations and activity of beneficial microorganisms are high, pathogens experience much difficulty in surviving. However, cropping schemes by the farmer impact two major factors which contribute to build-up of pathogens:
- Concentrated density of host tissue which provides opportunity for infection, survival, and proliferation.
- Gradual to sudden reductions in beneficial microbiology rendered through various cultural operations.
The resulting outcome is an increase in pathogen populations coinciding with reductions in competition, all of which add up to an unnaturally high density of pathogen inoculum (the part of a pathogen which causes infection) and concomitant pressures.
Why is it that healthy plants appear to resist disease more successfully than unhealthy plants?
All plants host a degree of resistance [R] to disease. Thus, in the great majority of cases a resistance response on the part of the plant represents an “expression of Inherent Resistance.” The degree to which this expression gains momentum over the opposing forces of the pathogen will govern whether or not infection and establishment of the pathogen ensues. Because the resistance response is linked to the “Rate of Metabolism,” we may define the R response as a “Rate-Related Response.” This R response is literally a race between the host and pathogen. The host attempts to produce a number of defense metabolites (warding off compounds) while the pathogen seeks entrance. This R response is best defined in graphing the basic differences between a susceptible versus resistance plant:
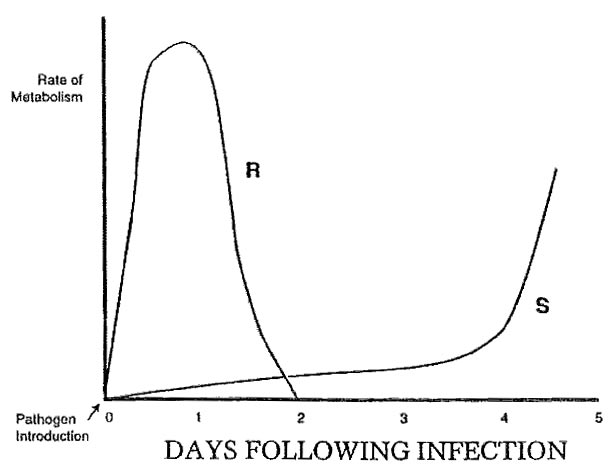 Graphic representation of the basic difference in metabolism between the susceptible plant response (S) of an unhealthy plant specimen versus the resistant plant response (R) of a healthy plant specimen. Upon attempted entrance of the pathogen, the S delays its response and production of walling-off compounds, while the R plant counters with a rapid rise in metabolism and production of walling-off compounds.
Graphic representation of the basic difference in metabolism between the susceptible plant response (S) of an unhealthy plant specimen versus the resistant plant response (R) of a healthy plant specimen. Upon attempted entrance of the pathogen, the S delays its response and production of walling-off compounds, while the R plant counters with a rapid rise in metabolism and production of walling-off compounds.
Please note that only a healthy plant would be able to respond quickly in a manner referred to as the R response. A plant with a compromised metabolism will respond slowly. By the time it does initiate heightened metabolism, the pathogen has invaded and established. Additionally, a healthy plant is generally endowed with tissue of higher integrity over that of the unhealthy counterpart, the order of difference analogous to cardboard versus tissue paper.
Why do most pathogens gain entrance to plants during rainstorms?
There are multiple factors at play:
- Adequate moisture to facilitate germination of a spore or reactivation of a bacterium.
- Predisposing events which weaken the host, such as:
- Reductions in photosynthesis but correspondingly minor reductions in respiratory- or energy-utilizing processes,
- Leaching of minerals by the distilled water-like rainfall,
- Minor to severe reductions in available oxygen for root metabolism and functioning,
- And occasional additions from rain and mobilization of existing nitrogen sources into the plant, thus requiring additional energy of assimilation (energy is diverted to the purpose of absorbing and processing nitrogen, leaving less energy to fuel an immune response).
The predisposing events basically act together to reduce the timely R response, with all conditions shifting to favor entrance and establishment of the pathogen.
Why are some areas notorious for plant diseases, and other areas relatively free of them?
Any differences in the frequency of plant diseases points to two main factors:
- The climate or microclimate of the region is more conducive to disease.
- The region hosts a heavier burden of disease inoculum (organisms or parts of organisms which cause infection).
While the first parameter relates to temperature and moisture factors, the latter becomes a bit more complex. For one, local inoculum sources may be heavier (e.g. abandoned orchards or vineyards). Secondly, the specific region may be within a “positive eddying zone” whereby the swirling wind currents concentrate particulates, including windborne inoculum.
This latter factor, for example, is of common occurrence for areas situated astride the foothills, off of which wind currents swirl and provide a means of concentrating disease inoculum. In California, some areas of Reedley, Kingsburg, and Arvin are prime examples of high disease pressure regions:
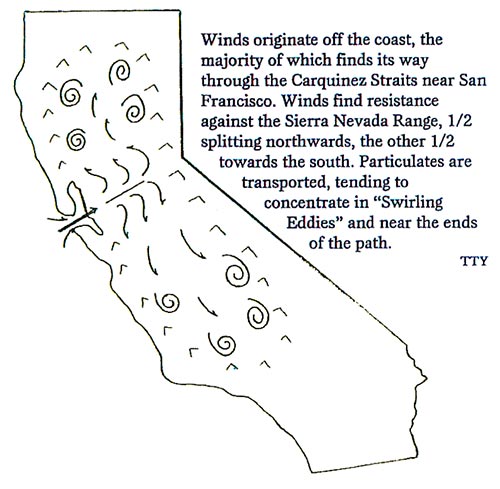 How does fallowing and/or crop rotation reduce pathogen populations?
How does fallowing and/or crop rotation reduce pathogen populations?
Absence of a host under fallow treatment of the land minimizes opportunity for reproduction. Further, the refraining from discing, plowing, concentrated fertilizer applications and general disturbance of the soil allows the gradual buildup of beneficial soil microbiology. Some of the microbial species are directly antagonistic while others render indirect natural suppression of plant pathogens. Additionally, the diverse ecology of microorganisms work concertedly to develop growth parameters of the soil. The superior growth parameters, then, contribute to expressing inherent resistance.
What is the connection between early planting and greater incidence of disease?
The primary connection resides in early plantings being subjected to cool and/or wet soils and concomitantly reduced growth rates of the host plant. As we discussed above with regard to why healthy plants are more disease-resistant than unhealthy plants, the ability to generate high rates of metabolism is necessary in order to resist pathogen invasion. Biochemical reactions are especially influenced by temperature fluctuations, the rate of metabolism generally rising with increased temperature.
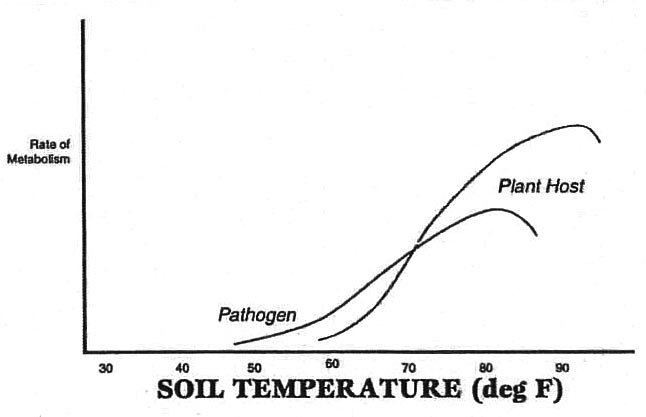 Graphic depiction for the metabolic activity curve of a pathogen versus its host. By waiting until soil temperatures reach 65 degrees Fahrenheit, for example, we will be able to minimize the degree of metabolic advantage the pathogen holds over that of the host, thereby minimizing opportunity for infection.
Graphic depiction for the metabolic activity curve of a pathogen versus its host. By waiting until soil temperatures reach 65 degrees Fahrenheit, for example, we will be able to minimize the degree of metabolic advantage the pathogen holds over that of the host, thereby minimizing opportunity for infection.
How do most pathogens get into a planted field?
Depending on the pathogen and specifics of the host environment, there are a number of avenues:
- Wind Transport – e.g. brown rot, powdery mildew, rust, grey mould
- Arthropods & Animals – e.g. anthracnose, walnut blight, fire blight, hull rot
- Irrigation & Flooding – e.g. Phytophthora, Fusarium, Verticillium, nematodes
- Nursery Stock – e.g. crown gall, stunt virus, Phytophthora, nematodes
- Seed – e.g. bacterial canker of tomato, Fusarium, black rot of crucifers
- Tools & Equipment – e.g. bacterial canker of stone fruit, fire blight, crown gall
You have emphasized the importance of soil microbial activity as an essential component of natural means to suppress and/or control soil-borne diseases. Why, then, do I still detect moderate to high inoculum levels and observe disease, despite the large volume of compost applied to my fields?
Enhancement of soil microbiology to achieve suppression and control of soil-borne pathogens within a short period of time (e.g. within the same season) requires that minimum levels of microbial activity are met. That is, a light to moderate rise in microbial activity and/or a gradual rise in microbial activity is incapable imposing sufficient natural suppressiveness to achieve pathogen control. Rather, it is both:
- A sudden rise in microbial activity.
- A high level of microbial activity that acts to severely encumber the pathogen populations with substantial pressures in space and time towards achieving more immediate suppression and control.
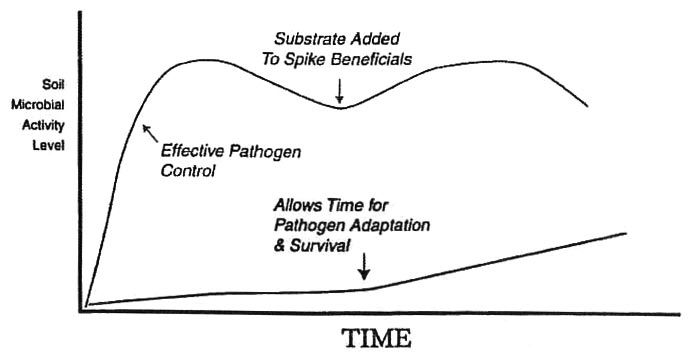 Level of microbial activity required to achieve pathogen control. Notice how a gradual increase in microbial activity is much too slow and with insufficient natural suppression, which then allows pathogen to either adapt and/or continue to populate.
Level of microbial activity required to achieve pathogen control. Notice how a gradual increase in microbial activity is much too slow and with insufficient natural suppression, which then allows pathogen to either adapt and/or continue to populate.
Notice how it is important to achieve an unnaturally high microbial activity (UHMA) immediately. As mentioned earlier, most pathogens represent varying degrees of specialization of the ancestral beneficial organism. One of the key principles in developmental biology states that as an ancestral organism responds to its environment with concomitant variations of form and physiology, the natural development is a shift from the general to the specific, a shift that can also be defined as one of the multiple adaptability to specificity, and thus the partial to total loss of versatility beyond its new, specific realm. Thus, in general soil-borne pathogens have adapted their form and physiology to enter the realm of the host, thereby minimizing a degree of competition. However, in so doing, the pathogen generally loses a small or large degree of fitness for the environment. When that environment is shifted to one of UHMA, with little opportunity for adaptation and adjustment, then, the pathogen is more likely to succumb to natural suppressive pressures.
 A panoramic, generalized view at the levels of microorganisms, their genetic makeup, and ability to adapt to varying environmental stresses. As most organisms become specialized, their adaptability to multiple stresses and variable competition (natural suppressive pressures) subsides.
A panoramic, generalized view at the levels of microorganisms, their genetic makeup, and ability to adapt to varying environmental stresses. As most organisms become specialized, their adaptability to multiple stresses and variable competition (natural suppressive pressures) subsides.
The phenomenon of UHMA can be quantified by various types of tests. Below are some guidelines that may serve as a benchmark:

(Thumbnail source: Max Pixels, public domain via CC0)
